


Thylacosmilus: The Southern Sabre


Pliocene South America was a strange place. Isolated as an island continent since the end of the Mesozoic, it developed a unique biota. While eutherian mammals- placentals- dominated the northern hemisphere, here they coexisted with metatherians- all mammals more closely related to marsupials. Additionally, the placental lines here were widely divergent. While the rest of the world was dominated by Boreoeutherians- that’s everything from mice and men to dogs and horses- with a healthy dose of elephantine Afrotherians, in South America, the Xenarthrans ruled the roost alongside the Meridiungulates, a group of hoofed mammals with uncertain affinities. Here, rhino-like toxodonts and camel-esque litopterns roamed in vast herds across the continent, alongside glyptodonts and giant ground sloths.
Hunting these creatures were not wolves, bears, or felines- again, these did not exist yet in South America. Instead, the apex predators of the continent were the “terror birds”. Formally known as Phorusrhacids, these were swift and aggressive predators, more like their dinosaur ancestors than modern birds, that killed their prey by hammering it to death with their immense, heavy skulls.
Soaring high above the landscape, the teratorns were also formidable avian predators. These were the largest birds to ever fly. The largest, Argentavis, stood 5’11 on the ground and had a wingspan of 21 feet. Despite superficially resembling vultures, thanks to their eagle-like beaks they were likely active predators, in addition to filling a scavenging niche. With their impressive height and wingspans, teratorns would have easily intimidated most predators away from their kills, buying the giant birds a free lunch.

Not to be outdone, mammals offered some predators of their own. There was a giant carnivorous armadillo, Macroeuphractus, and the omnivorous, bear-sized Chapalmalania, which was most closely related to raccoons. But the most fearsome mammalian predators were the sparassodonts. These were metatherians, closely related to marsupials but phylogenetically just outside that group. They converged strongly with placental predators in many different niches. Some, like Proborhyaena, were the size of bears, while others like Cladosictis were more like large weasels. The creature we’re discussing here, Thylacosmilus, convergently evolved to fulfill the same niche as sabretoothed cats.
The sparassodonts were limited by competition from the terror birds. They couldn’t compete with the speedy phorusrhacids on the open plains, so they were largely restricted to dense forest biomes. This was, eventually, to be their downfall. In the west, the Andean orogenesis was well underway, with the newborn mountains gradually creating a vast rainshadow and drying out the interior of the continent. After flourishing in the Paleocene and Eocene, the sparassodonts began to decline after the mid-Miocene. By the end of the Pliocene, 3 million years ago, only one sparassodont family remained, and it is to this last group that Thylacosmilus belonged.

The “Marsupial” Sabretooth
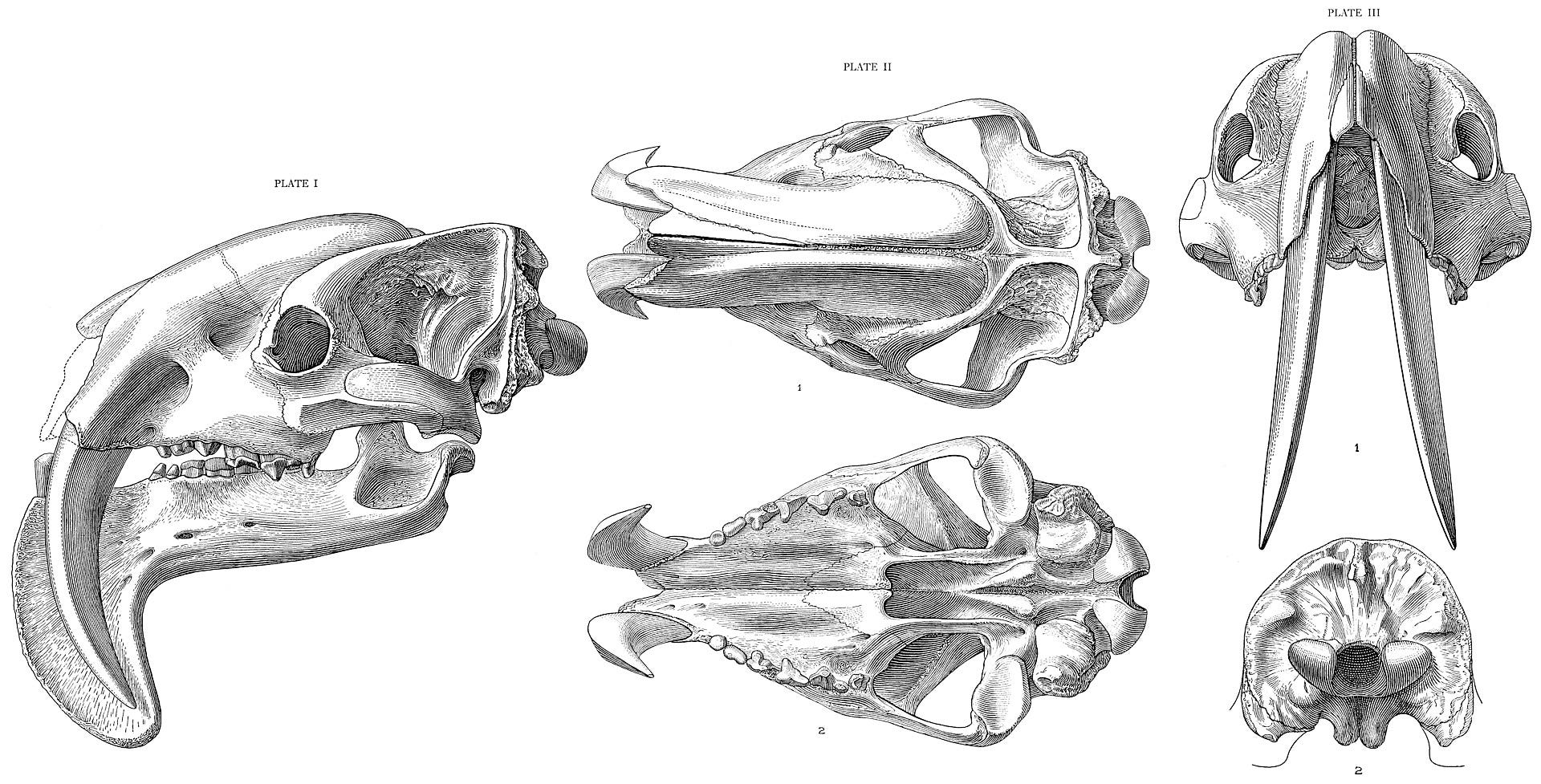
Thylacosmilus atrox was discovered in 1926 during the second Marshall Field Paleontological Expedition, in the Catamarca Province of northern Argentina. It wasn’t properly described until 1933, when it was christened by American paleontologist Elmer S. Riggs. The name means “cruel pouch knife”, reflecting the animal’s marsupial affinities and its sabre canines. Riggs knew that Thylacosmilus was related to known members of the sparassodont family, but he also recognized that that it was highly specialized, and so he placed it in its own subgroup- the thylacosmilids.
The thylacosmilids spring onto the fossil record near fully-formed, leaving their evolutionary origins a mystery. There are only four known thylacosmilid genera. Thylacosmilus was, of course, the first to be described, and no other genera were identified until the mid-Miocene aged Anachlysictis was discovered in 1997. This genera was more primitive than Thylacosmilus, and only weighed about 40lbs, but was still well along the sabretooth path. Although its dentition more closely resembled basal sparassodonts than its descendant, it already possessed sabre teeth.
To quote Riggs’s paper, Thylacosmilus was “one of the most unique flesh-eating mammals of all times.” Unique it certainly was, yet it bore a strong superficial resemblance to placental sabretooth cats, thanks to convergent evolution.
Weight estimates for Thylacosmilus vary, because there isn’t really a one-to-one comparison between sparassodonts and other carnivorous mammals. Extant carnivorans walk differently (thus their limbs support their weight differently), extinct marsupial predators tended to be lightweights, and the sparassodonts themselves had large heads relative to their bodies. All of this together makes overestimation an issue.
Nevertheless, Ercoli and Prevosti (2011) came to a weight range between 180 and 260lbs, based on Thylacosmilus’s limb proportions. Since the limbs directly support the animal’s weight, their proportions are ideal for inferring an animal’s mass, though in this instance with a wide range to account for the uncertainties of sparassodont locomotion. Sorkin (2008) came to a higher estimate of 330lbs, based on the animal’s skull proportions. Assuming this estimate as a high-end, we can infer that Thylacosmilus weighed on average about the same as a jaguar, with large individuals potentially weighing as much as a lioness. This would make Thylacosmilus one of the largest carnivorous metatherians known.

The limbs of Thylacosmilus were stocky and robust. The hind legs- with a thick femur, short tibia, and semi-plantigrade feet- suggest that Thylacosmilus was an ambush predator, as do its rigid lumbar vertebrae. The flexibility of the femur head also suggests that the animal was able to stand on its hind legs, which would have been very helpful for prey capture, enabling it to grapple with prey and pull it to the ground while standing upright. Ercoli et al (2012) also suggests that it may have been capable of some degree of cursoriality- not sprinting, but a kind of long-distance, “persistent galloping” similar to hyenas and certain species of bears.
At the forelimbs, the humerus supported enormously strong pectoral and deltoid muscles. This was crucial for the animal’s hunting style. Sabretooths like Thylacosmilus needed their strong forelimb musculature to knock down prey and, perhaps more importantly, to restrain it for the killing bite. Prey restraint was critical to the sabretooth lifestyle- an unrestrained victim thrashing around could easily have broken the predator’s fragile canine teeth.
The lower humerus also supported strong supinator muscles, which would have given the animal flexible hands, able to pronate, supinate, and laterally rotate. This dexterity would also have been vital for grappling with and restraining prey. Every one of these traits was also present in placental sabretooths like Smilodon, and was acquired by Thylacosmilus independently via convergent evolution.

One major departure from its convergence with felines is that Thylacosmilus lacked retractable claws. This has been taken by some researchers to mean that it wasn’t actually filling in a sabretooth niche, since the dexterity offered by retractable claws was vital to felid and para-felid sabretooths. Because Thylacosmilus didn’t have them, it must have been filling some other niche that didn’t involve dynamic prey capture. This simply doesn’t hold up to scrutiny; bears also lack retractable claws, but are nevertheless quite capable of pinning large prey.
Instead, Thylacosmilus’s lack of retractable claws appears to be more due to pedigree. While retractable claws are an ancestral trait in placental carnivores, the sparassodonts and South American marsupials don’t show any evidence of ever having possessed them. To compensate for this apparent handicap, Thylacosmilus instead had extremely broad and flexible paws, and partly opposable thumbs. These adaptations would have helped Thylacosmilus excel at wrestling with and pinning down prey, despite lacking a major tool from the placental arsenal. It’s a good example of how evolution can work with divergent forms and use different methods to achieve the same results- in this case, enabling an ambush predator to take down its prey.

Thylacosmilus’s neck was very strong. The cervical vertebrae have prominent transverse processes and ventral keels, which are associated with powerful longus colli muscles, crucial for flexing the neck. There are also insertion points on the skull for the longus capitis, another powerful flexor which is a continuation of the longus colli, which would have given Thylacosmilus both immense strength and delicate control over its head motions (Anton 2013). This will all be important in a moment when we discuss how Thylacosmilus killed its prey.
Related to the neck, the skull occiput is raised in Thylacosmilus. This condition is also present in several unrelated sabretooth lineages, including the machairodonts and barbourofelids, suggesting that it was an important adaptation for the sabretooth lifestyle. Most carnivores have nearly horizontal occiputs. The higher occiput of sabretooths would have enabled them to better raise their whole heads relative to their necks. This makes sense, given the very wide jaw-gapes of sabretooths- and as we shall see, Thylacosmilus’s was the widest of them all- because it would help open the mouth as wide as possible just before biting, and give the head a wide arc to work with when the mouth was suddenly closed in the downward thrust of the killing bite. (Anton 2013)
You probably have noticed from the artwork that Thylacosmilus had a very prominent chin, tracking downward vertically with the path of the canines. This structure is called the symphyseal flange, and it served two purposes. The first was to protect the canines against unnecessary lateral stresses- when it was grappling with prey, if it got kicked in the mouth the pronounced chin would protect the fragile sabres somewhat. Additionally, it would have helped the mandible withstand the strong vertical forces of the killing bite. However, like the sabres themselves, the flange would have been vulnerable to lateral stress, which would have required Thylacosmilus to keep its prey as immobile as possible.

Overall, Thylacosmilus’s skull was quite robust. Goin and Pascual (1987) posit that it may have held its head lower than other animals, simply because the massive canines occupied the entire front of the skull and may have impaired the animal’s vision if it held its head level. If it did indeed hold its head with a greater natural bend, then the dramatic symphyseal flange would have served the additional purpose of protecting the animal’s throat from its own canines.
They also suggest that, assuming Thylacosmilus did hold its head at a dramatically low angle, it might have actually headbutted its prey to the ground, rather than using its paws to grapple with prey like a typical sabretooth. Its already robust skull would have been naturally reinforced by the canines, which grew in an arc over the entire maxilla, giving it a distinctly convex shape, like a football helmet. This would square with Riggs’s initial speculation about the network of furrows and pits on the upper maxilla hosting a keratinous “horn covering” which possibly could have aided in headbutting.
There are, however, good reasons to doubt this theory. Firstly, as Goin and Pascual admit, it would be “unique among living and fossil vertebrates” for a predator to capture prey via headbutting. Sometimes there really are new things under the sun, but such a bizarre prey capture method would still likely have evolved more than once if it was effective. And it probably wouldn’t have been very effective, for the simple reason that it would have severely limited Thylacosmilus’s means of catching dinner. It would have to make perfect straight-line headbutts at its prey to have a hope of knocking it down, assuming it could even put that much kinetic energy into the hit without hurting or killing itself.
Animals that engage in headbutting behavior, like deer and bighorn sheep, usually have antlers and horns to bear most of the force of the impacts. Thylacosmilus would have to ram prey with its forehead. The closest analogue to this would probably be the tapinocephalians, a group of bizarre Permian therapsids which are known to have headbutted each other directly, no wimpy antler-cushions involved. We know they did this, because their skulls were extraordinarily thick- up to 4 inches at their thickest point- to protect their brains from the force of impact. They made Thylacosmilus’s robust head look fragile in comparison. Tapinocephalians likewise had very thick cervical vertebrae to brace their spinal cords. In contrast, Thylacosmilus’s cervicals, while having attachment points for very strong muscles, do not have any features suggesting they were capable of withstanding severe impact trauma.
So while we can safely discard the headbutting theory, it remains a mystery what exactly Thylacosmilus would have used its bizarre keratin horn for. If it was in fact a horn- there’s also a theory those pits and grooves on the skull were supporting some sort of soft-tissue structure, the purpose of which would also still be a mystery.

Thanks to its well-preserved skull, Quiroga (1988) was able to analyze the animal’s endocast- a natural cast of the cranial cavity, which can tell us the location of certain lobes of the brain and their sizes. Because of this, we know Thylacosmilus wasn’t very smart, similar to modern marsupials. Indeed, comparing Thylacosmilus to a horse-like litoptern that it coexisted with, it appears Thylacosmilus may have been less intelligent than some of its prey animals, with only half the encephalization and a quarter of the cortical area of the litoptern. As far as its somatic nervous system- the system governing voluntary muscle control- 18% was devoted to eyesight, 7% to hearing, and 42% to the oral and mandibular regions. Control of its jaws was obviously quite important.
Additionally, Thylacosmilus had very small eyes, and limited stereoscopic vision, with an overlap of a mere 50-60°. This is pitiful compared to living predators like lions and wolves, but isn’t actually that far off-base from other sabretooths like Smilodon, although its vision was still better. Placental sabretooths generally seem to have relied more on their acute hearing than vision to hunt their prey. However, given that Thylacosmilus devoted more of its brain to eyesight than hearing, it seems like it was primarily using its poor eyes to hunt.
I mentioned jaw-gape earlier. Thylacosmilus easily had one of the most dramatic jaw-gape of any known animal, capable of opening its mouth a full 105.8°. This is laughably wide, much wider than the gape of the famous sabretoothed cat Smilodon (87.1°) or the leopard (72.6°). It also means, counterintuitively, that Thylacosmilus had absolutely no chance of killing prey with its jaws alone. The wide gape did not make the jaws more effective at biting- jaw-driven muscle bite force in Thylacosmilus has been calculated at a paltry 38 Newtons, extremely weak and certainly not enough to make efficient use of the sabres. Additionally, skeletal stress distribution models suggest that Thylacosmilus would probably have shattered its jaws at the hinges if it tried to kill prey using bite force alone- this is also the case for Smilodon. Instead, the wide gape was so that the sabres could be brought to bear by the head depressors and cervical musculature. It killed with its neck muscles, not its jaw muscles.

The Cruel Knives
This brings us to the business end of the animal. Thylacosmilus probably had the most specialized dentition of any sabretooth mammal to ever exist, representing a complete commitment to that lifestyle. It’s important for the reader to understand this- sabreteeth are not something an animal simply adds to its armory. The elongated canines are quite fragile, and require an entirely different suite of adaptations to be effective. They aren’t simply stabbing weapons, and have to be deployed with great precision against specific parts of the body, or else they’ll simply shatter against the bones of their prey.
Sabretooth predators had a specialized method of killing called the canine shear-bite. “Canine” here referring to the canine teeth, not dogs. A sabretooth’s killing bite began with the predator opening its jaws to full gape against a convex portion of the prey’s body, like the throat. As stated, the jaw muscles were too weak to exert enough force to get the canines to penetrate the flesh, so upon contact the head depressor muscles went into play, pulling the skull down like a lever and sinking the canines in. The very strong neck musculature likewise went into action, providing more thrust from behind the skull to sink the canines in even deeper. This is why the animal’s neck musculature is so important, because those muscles are actually the key to the sabretooth kill, not the jaw muscles. The lower jaw serves only as an anchor on the prey’s body against which the head depressors act, until the fangs have sunk in to a sufficient degree that the jaw muscles can be brought to bear to close the bite. Thus, the prey would suffocate or die from massive blood loss with the sabres slicing through many vital arteries in the neck. (Anton 2013)

Thylacosmilus’s canines had extremely deep alveoli, with the roots arcing over the entire maxilla, up over the eye orbits, and terminating over the animal’s braincase. More than half of the tooth was hidden inside the skull, continuously growing throughout the animal’s lifetime. This was actually a marvelous adaptation. Sabreteeth are very fragile and prone to breaking under stress. Many feline sabretooth skulls have been found with broken sabres, so continuous growth would have enabled Thylacosmilus to- after a long enough time- heal from damage that would have been permanent for a placental sabretooth like Smilodon. The sabres were also self-sharpening, thanks to wear against the lower canines.
One thing Thylacosmilus lacked was a strong incisor battery. All other known sabretooths- machairodonts, the non-feline nimravids and barbourofelids, and even the Permian gorgonopsians- had strong incisors between their sabres to help anchor their jaws onto their prey. It’s unclear if Thylacosmilus even had upper incisors- in all known specimens the premaxilla is broken or missing, and the space between the sabres was very narrow anyway. They likely did have small incisors, due to wear on the lower canines matching with the presence of upper incisors, but regardless these would have been virtually useless for prey capture. The lower canine teeth were likewise reduced, but still functional. Again, it appears Thylacosmilus was near-fully optimized for the sabretooth lifestyle, not even needing incisors to kill thanks to its immense canines. Thylacosmilus’s canines were shaped differently than any other sabretooth, slender and triangular in cross-section, as opposed to the oval shape of placental sabres. These triangular canines may have actually been more effective at penetrating flesh, thus eliminating the need for anchoring incisors.
Another unusual dental adaptation is the cheek teeth. In placental carnivorans, the upper fourth premolar and lower first molar are modified into carnassials. These blade-like teeth are paired together and act as shears for cutting meat. If you have a dog, you’ll easily see his carnassials when he pants. Now, metatherians don’t have true carnassials (although some did later acquire a similar dental plan), so instead, Thylacosmilus modified all of its post-canine teeth into a continuous shearing blade. This is, quoting Mauricio Anton’s excellent book Sabretooths: “the most extreme adaptation to hypercarnivory among South American marsupials.”

Sabretooth or Scavenger?
There is, however, an alternative theory, that Thylacosmilus wasn’t much of a predator at all. Janis et al (2020) proposed that Thylacosmilus was not, in fact, a metatherian equivalent to sabretooths like Smilodon, but rather an obligate scavenger. They noted that the shape of the mandible, the lack of upper incisors, the incredibly weak bite-force, and the somewhat splayed position of the canines point to Thylacosmilus being unsuited for the high precision canine shear-bite of other sabretooths.
They suggest that the “sabres” were in fact more akin to tusks, and that Thylacosmilus’s diet was mostly soft foods. Specifically, they propose that it was an intestine specialist, using its massive jaw-gape and sabres only to open fresh cacarcasses, using its incisor-less premaxilla- which is presumed to have had fleshy lips like a walrus- and muscular tongue to suck the internal organs out, more specifically the intestines.
That’s ridiculous.

First of all, we’ve already discussed how Thylacosmilus likely did have small incisors, due to wear on the lower canines, but that they weren’t necessary for the killing bite because its canines were actually more efficient at penetrating flesh than those of other sabretooths. Secondly, the paper fails to address why the sabres being used to slice open carcasses would leave zero evidence of such wear. They also suggested that the cheek teeth- which, recall, were modified into a single shearing surface- actually show no signs of shearing wear, and while this may suggest Thylacosmilus was chewing in a different way than placental sabretooths and other sparassodonts, it certainly does not point to Thylacosmilus being an obligate scavenger.
Parenthetically, Domingo et al (2020) analyzed the carbon isotopes present in Thylacosmilus’s teeth and found that its diet mainly consisted of C3 plant-consuming animals- large caviomorph rodents and notoungulates, the exact animals it would have been hunting if it were an active predator.
Finally, and this is the most important line of evidence to debunk this silly theory, is that there is no such thing as a terrestrial obligate scavenger. They don’t exist, because there is simply never enough carrion on the ground to sustain that lifestyle. If you ever go to the Serengeti, you’re not likely to see many dead animals. Instead, you’ll see lots and lots of living ones, because living things like to stay alive. They will all die someday, of course, but not at a rate sufficient to permit an animal to eat nothing but carrion. Only vultures, which can effortlessly glide over vast territories, wheeling around and around on the thermals in their endless search for carrion, are able to get by on a purely scavenging lifestyle.
Ah, but what about hyenas? Hyenas, commonly misconceived as obligate scavengers, are in reality active predators. Even the brown and striped hyenas, which are primarily scavengers, still supplement a significant portion of their diets with their own kills, and also with fruit and eggs. And they certainly aren’t specialists in soft tissues. Rather, hyenas are excellent bone-crushers, with powerful jaws and giant premolars designed for this purpose. When you’re adapted to scavenging, you take whatever part of the carcass you can carry, because other, bigger predators will come to enjoy the free lunch too.
In an environment populated by giant carnivorous birds, Thylacosmilus simply wouldn’t have had time to leisurely slurp up the soft tissues of a dead animal. The terror birds would have arrived and kicked it to death in short order, or teratorns would have swooped down to scare it off. Additionally, if Thylacosmilus were in fact an obligate scavenger, it would had to have been bullying these giant birds away from their kills, as modern hyenas do to leopards and wild dogs. This is simply not a plausible scenario, the birds were way bigger and more aggressive.
Relatedly, predators were thin on the ground in Pliocene South America. The phorusrhacids, teratorns, and the handful of mammalian predators would have been the only providers of carrion outside of natural causes. There simply wouldn’t have been enough carcasses to sustain a whole population of Thylacosmilus. In short, it was perfectly well adapted to the sabretooth lifestyle and this study is terribly wrong. Melchionna et al (2021) does rescue the study somewhat, suggesting Thylacosmilus was still an active sabretooth predator, but simply preferred to dine on the soft internal organs of its victims rather than their flesh, which fits the behavior of living predators.
The End of the Sabre

The final extinction of Thylacosmilus has been a subject of debate. Traditionally, it was assumed to have gone extinct after GABI- the Great American Biotic Interchange, which occurred when the Isthmus of Panama connected North and South America for the first time since the Mesozoic. The story goes that after Smilodon entered South America, it competed with Thylacosmilus for food resources, and in the dramatic battle of sabre-versus-sabre, Thylacosmilus lost and went extinct.
While it’s a great story, the timeline unfortunately doesn’t add up. Firstly, Thylacosmilus appears to have gone extinct during the Piacenzian epoch of the late Pliocene, between 3.6 and 2.5 million years ago. This is right on the cusp of GABI, which is believed to have begun about 2.7 million years ago. But Smilodon is unknown in South America before the middle Pleistocene, a mere 781,000 years ago. There simply wasn’t an abrupt replacement of Thylacosmilus by Smilodon. Their respective existences never overlapped.
The discrepancy could be explained somewhat, however, by the Signor-Lipps effect. Since the fossil record is incomplete, the first and last individuals of a species will never be preserved as fossils. Thus, the youngest known fossils of a species don’t actually represent the very last one, and the extinction date presumed from the youngest known individual is only approximate. Sometimes we can pin down extinction dates precisely despite the gaps in our knowledge- for example, we don’t have any bones of dinosaurs that were actually killed by the Chicxulub asteroid. But for most genera and species, the extinction date is only proximal, and sometimes organisms can surprise us by being absent from the fossil record for huge stretches of time and then suddenly appearing alive and well in a more recent era. So, hypothetically, Thylacosmilus could have lasted well into the Pleistocene, and just didn’t leave a good fossil record for us to follow. However, given the overall decline of sparassodonts prior to GABI, it seems more likely that Thylacosmilus went out a different way.
Currently, the favored theory for Thylacosmilus’s extinction is climate change-induced habitat destruction. During the Miocene and early Pliocene, South America was covered in vast, dense forests, but this changed near the end of the Pliocene. In this part of South America, the Andes had been patiently growing since the Maastrichtian, their gradual rise punctuated by quick spurts of uplift. Their ponderous rise eventually created a rain shadow over eastern South America which grew with the newborn mountains. There was also a drop in global temperatures around this time, combining with the mountain rain shadow to aridify the Pampas.
The lush forests withered, first giving way to open woodlands, then to savannahs and thin strips of woodland lining the rivers and streams that crisscrossed the region. Finally, even these attenuated gallery woods disappeared, leaving us with the modern Pampas grasslands. While this is good for humans, creating a broad plain of fertile land, it was a disaster for Thylacosmilus. Dependent on the cover offered by the gallery woods to mount its ambushes, it couldn’t adapt to the changing climate.
Thus, by the time Smilodon walked across the Isthmus of Panama, one and a half million years later, it found itself in a continent empty of sabretooth predators, and it was perfectly adapted to fill in the niche left by the lost sparassodont.

Sources
Sabertooth by Mauricio Anton, 2013
An eye for a tooth: Thylacosmilus was not a marsupial “saber-tooth predator”, Janis et al (2020)
Nice reddit comment breaking down the above study and some of the counterarguments
A biomechanical constraint on body mass in terrestrial mammalian predators, Sorkin (2008)
Preliminary description of a new marsupial sabretooth from the Pliocene of Argentina, Riggs (1933)