




It is the Albian epoch, a hundred and thirteen million years ago. The primordial supercontinent Gondwana is fracturing, its cratons awakening from ancient slumbers and beginning the long drift towards their geologic destinies. India, Africa, and South America the granite shards of a broken land. The lost continent Zealandia begins to cleave off from the east, doomed to slowly sink into the South Pacific. Australia alone remains joined to Antarctica, but even this inconceivably ancient connection is beginning to fray. What will eventually become the Southern Ocean is at this time a great rift valley, the Australian Bight beginning to form as a narrow bay between the Nullarbor and Wilkes Land. The Queensland coast, pocked by cinder cones and fissure vents, has been erupting for ten million years, and it will continue for ten million more. The weeping scablands are scoured by sheetfloods that carry ash and pumice all the way to Victoria, nearly 1,700 miles away, where it fertilizes the floodplains of the broad, braided rivers that wind through the region, allowing vast wealds of lush temperate rainforests to spring up. The Perth, Canning, and Great Artesian basins, someday destined to become parched red outback, are presently submerged beneath iceglazed inland seas.
The entire eastern half of Australia lies deep within the Antarctic Circle, yet there are no permanent ice caps. The average temperature at the south pole dances between 21-46°F, occasionally dipping down into the teens and the aughts as the climate fluctuates. Glaciers are molded and melted every few centuries, the icy pulse of a greenhouse world. In summer, the midnight sun dances along the horizon for three months, before finally exiting the stage and leaving a warm outro of twilight to last through autumn. Then the crepuscular afterglow fades, and the months of darkness begin. Flurries dusting the ferns and ashblack soil, gradually changing into howling snow squalls as the long polar night settles in. Each flake a white orison to the vanished sun, answered only by the silent auroras shimmering overhead.
It is into this world that Leaellynasaura was born.

Leaellyn’s Lizard
Leaellynasaura amicagraphica was a very small ornithopod dinosaur, standing at a mere one foot four inches. It lived during the late Aptian and early Albian epochs of the early Cretaceous, between 118 and 110 million years ago. It’s only known from fragmentary remains, so classifying it has been a nightmare. Paleontologists know it was an ornithopod- the clade of dinosaurs including famous “duck-bills” like Hadrosaurus and Parasaurolophus, as well as dozens of larger and smaller genera- but getting much further than that is a challenge. Most often, Leaellynasaura has been classified as a “hypsilophodontid”, which is a wastebasket taxon where paleontologists have lumped together a bunch of small ornithopods whose phylogenetic relationships are uncertain. The most recent analysis, Herne et al 2019, placed it in the clade Elasmaria, a group of bipedal ornithopods that resided in Gondwana- what was left of Gondwana- during the Cretaceous. If you’ve ever seen a dinosaur movie or series where there’s a generic type of small, skittish bipedal dinosaur constantly fleeing from a big, hungry theropod, this is them. A rather boring, unassuming group of dinosaurs.
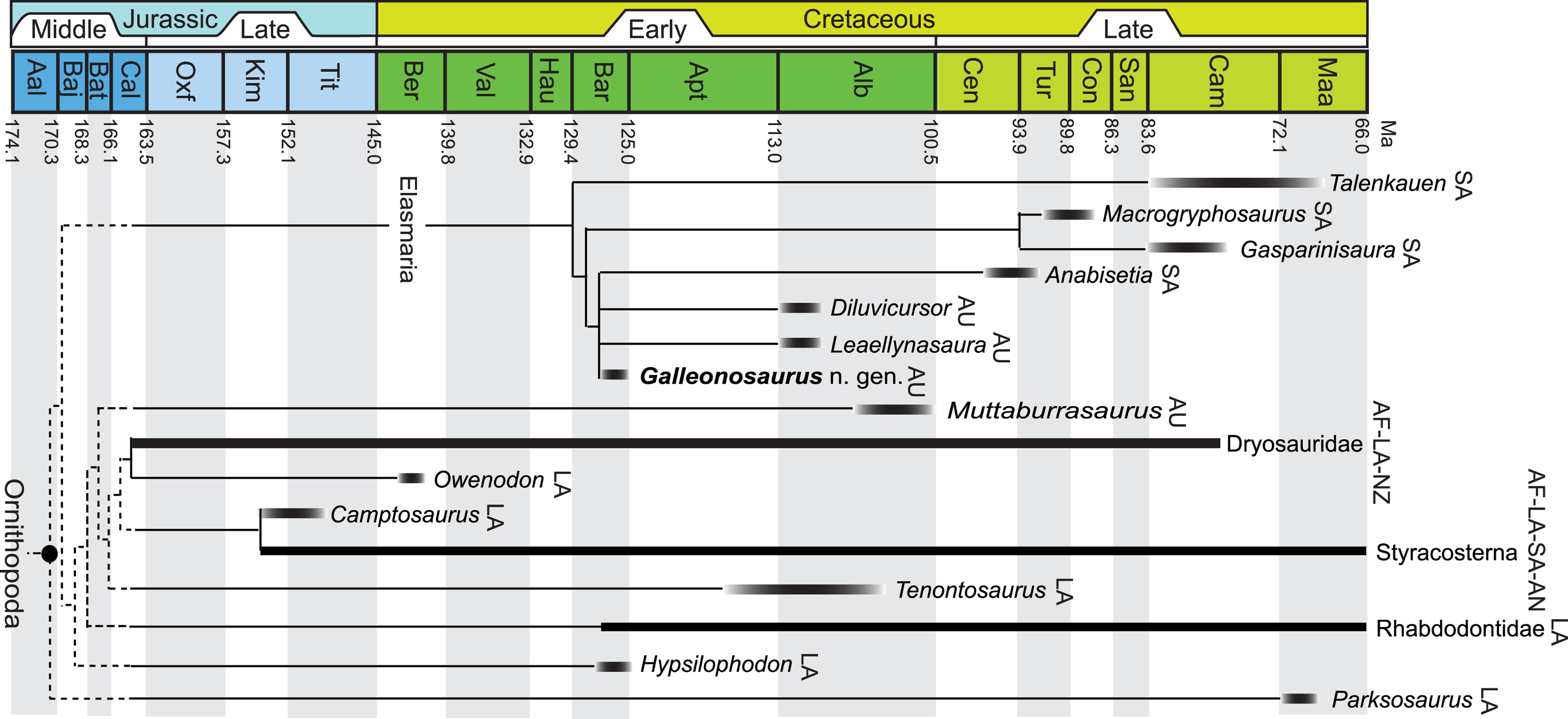
Leaellynasaura was unique though.
A word of warning before we begin- reconstructing Leaellynasaura will require a bit of detective work. As previously stated, it’s known only from very fragmentary remains, and paleontologists don’t even agree on whether or not all of these actually belong to the same species. Nevertheless, we will go in with the assumption that they do, and then pick it apart at the end.

Night Vision
Beginning at the head, Leaellynasaura had unusually large eyes for a dinosaur its size. Big saucer-eyes on the sides of the head, as would be expected for a small herbivore. Such large eyes would have provided excellent monocular vision for detecting predators on the animals’ flanks, and also would have gifted it with exceptional night vision, similar to an owl or a loris.
It’s also been suggested that the large eyes were simply due to the specimen- we only have one skull- being from a juvenile. The specimen was definitely a juvenile, about two-thirds its adult size, but it’s unknown whether or not the skull would have continued to grow and thus proportionally “shrink” the eyes.
A bit of speculation that often pops up in depictions of Leaellynasaura is portraying it with deep blue or purple eyes. It’s analogous to the eyes of modern reindeer, which change color with the seasons- in summer, their eyes are brown, but in winter they turn blue or purple. This adaptation allows them to see much better- their summer eyes reflect 95% of the light that hits them, but their violet winter eyes only reflect 40%. They can also see in the ultraviolet range, so things that would be invisible to the human eye against a backdrop of snow, like wolf urine or white fur, are sharply visible to a reindeer. Giving Leaellynasaura- or any dinosaur, for that matter- purple eyes is totally speculative, and there is no way we can ever find out what color a dinosaur’s eyes were. But it’s something that would make sense for Leaellynasaura to have, given its polar environment.

Regardless of whether or not the eyes were really proportionally larger than normal, a partial endocast from the same skull preserves an impression of the top of Leaellynasaura’s brain* and confirms that the species had exceptionally good vision, with two large nubs at the rear representing an enlarged optic lobe, a perfect complement to those giant eyes. The endocast is also very detailed, suggesting that Leaellynasaura had a large brain that filled nearly the entire cranial cavity.
*I wish to stress that an endocast is not a fossilized brain. It is a detailed impression of the brain left on the inner surface of the skull. Endocasts can show us specific lobes of the brain, and we can even take a whack at guessing an animal’s intelligence from endocasts, though the results are highly debatable.

The huge size of Leaellynasaura’s optic lobe- also known as the optic tectum- has implications for the animal’s behavior. In non-mammalian vertebrates, the optic tectum doesn’t just help to process visual stimuli. It’s also largely responsible for quick reactions to that visual stimuli. Frogs use their optic tectums to target insects with their tongues. Snakes use theirs to deliver bites with incredible precision. In birds, the optic tectum is often one of the largest parts of the brain, helping them to navigate the challenging, three-dimensional environment of the sky; pterosaurs likewise had large optic tectums for the same reason. In mammals, the equivalent brain structure is known as the superior colliculus, and it’s larger in arboreal mammals that also have to navigate the complex, three-dimensional spaces of forest canopies. Leaellynasaura couldn’t fly, but it may very well have been arboreal.
The arboreal theory was initially suggested way back in 1882 by fossil collector John Whittaker Hulke, talking about Leaellynasaura’s distant cousin Hypsilophodon being arboreal, or at least able to climb trees for safety thanks to their opposable thumbs. Some of the arguments made by Hulke and fellow paleontologist Othenio Abel were wrong- they thought Hypsilophodon’s first toe functioned as an opposable hallux, due to inaccurate musculoskeletal reconstructions- but the theory remains possible for other small ornithopods whose skeletons are more fragmentary. We don’t know if Leaellynasaura had opposable thumbs or toes. It might have. Unless we find more material, we won’t know.
It’s also possible that Leaellynasaura’s large optic tectum was helpful for procuring food. While it was herbivorous, it probably wouldn’t have turned its nose up at insects. Bugs are really easy to eat, even for animals with teeth designed to process plants. Insectivory isn’t that specialized- a single bite to crush the exoskeleton is all the processing required. And many animals conventionally thought of as herbivores will occasionally eat meat- for example, deer will sometimes snack on rabbits, bird eggs and hatchlings, and even cannibalize other dead deer. Similar behavior has been observed in cattle, squirrels, and tortoises. Despite being herbivores, sometimes their diet doesn’t give them all the protein they require, and bugs or hatchlings are a perfectly good source of it. Perhaps Leaellynasaura supplemented its herbivorous diet of ferns and horsetails with the bountiful supply of insects in the temperate rainforests it inhabited, in which case it would have needed the enhanced visual processing to help it catch fast-moving airborne targets. Or maybe the enlarged optic tectum was a result of some combination of these behaviors.
Tall Tail
Leaellynasaura also had a phenomenally long tail, the longest tail relative to body size of any dinosaur currently known. It had over 70 caudal vertebrae, making up three-quarters of the animal’s total body length.


Uniquely, the tail lacked ossified tendons. Ossified tendons are tendons that have turned into bone, fortifying the muscles in a given part of the body. In other ornithopods, these bonified tendons form a latticework along the lower back and tail, stiffening the tail and holding it straight out to help balance the animal. Leaellynasaura’s lack of these tendons would have given its tail incredible flexibility compared to any other ornithopod. This isn’t to say that the tail was a noodle. It still needed some structural stability to help balance the animal. It just used an alternative to ossified tendons that gave it a much greater range of motion.
The wonderful flexibility of Leaellynasaura’s tail has led to much speculation over what exactly it was using it for. The most common (and cutest) theory is that it used its tail as a kind of scarf, wrapping it entirely around its body for warmth, much like how the arctic fox will curl up into a ball and use its tail as a blanket. Recall that Leaellynasaura lived very close to the South Pole- it was warmer in the Cretaceous, but still pretty damn cold, so there would have been strong selection for traits that keep animals warm.


The adorable blanket-tail theory is largely contingent on Leaellynasaura possessing feathers. However, currently there’s no direct fossil evidence that it did. Feathering in ornithischians- the group of dinosaurs including ornithopods, ceratopsians, ankylosaurs, stegosaurs, and pachycephalosaurs- is scant compared to theropods. We know the larger hadrosaurs did not possess feathers, thanks to scale impressions from many different species, as well as a few excellently preserved Edmontosaurus “mummies” which preserve nearly the entire skin of the animal, showing they had nothing but small, non-overlapping scales. But at the same time, we know that some ornithischians did possess feathers. Two species of primitive ornithischians belonging to groups basal to ornithopods, Tianyulong and Kulindadromeus, are known to have had feathers, implying that feathers were originally present in all dinosaurs, and some lineages later lost theirs, while others retained them.
The fact that two ancestors of Leaellynasaura had feathers means that phylogenetic bracketing can be used to infer Leaellynasaura may have had feathers too- such a beneficial trait for retaining body heat would probably have been retained down the generations. Kulindadromeus specifically lived in Jurassic Siberia, 168 million years ago- exciting, because Siberia was in the Arctic even then, suggesting the feathers were used for retaining body heat.
Confusingly, the tail of Kulindadromeus appears to have been feather-free, but this doesn’t necessarily mean Leaellynasaura’s much longer tail was also unfeathered. If your only knowledge of mammalian fur came from a mouse or an opossum, it wouldn’t logically follow that every mammal’s tail was un-furry. Leaellynasaura having feathers is something that isn’t proven, but it’s another one of those things that would make sense, given the environment it was living in. It would make sense for the animal to have a warm, fluffy coat of feathers for insulation.
The tail may also have been used as a social or threat display. Many birds are able to raise their hackles- puff up their feathers- to make themselves look bigger and more intimidating, which is useful both for territorial displays to rivals, and for startling predators. This raises the possibility of some extremely fluffy Leaellynasaura tails, with the tail possibly appearing as large as the rest of the animal. In addition to its application as a display, being able to puff up the tail- and feathers on the rest of the body- may also have served the more practical purpose of simply keeping the animal warm. Owls warm themselves by puffing out their feathers and then lowering them to trap many small pockets of air around their bodies, which are warmed up by the bird’s own body heat. The whole process is just creating a blanket around the owl. It’d make perfect sense for a tiny polar dinosaur to have a similar adaptation, especially on its long, spindly tail which would have been very vulnerable to frostbite.

We can also let our imaginations run wild with thoughts of Leaellynasaura having a colorful, peacock-esque tail used for mating displays, or exploiting the tail’s flexibility for elaborate mating dances. Such a distinguishing feature would be perfect for sexual displays. Maybe longer tails, rather than being used as an effective threat display, were detrimental to Leaellynasaura’s chance of surviving predator encounters. Similar to how the magnificent tail plume of a peacock increases his visibility to predators and handicaps his ability to quickly get airborne to evade them. Only the fittest peacocks are able to maintain a large tail fan and survive encounters with predators, so a huge plume is an honest signal of a peacock’s fitness to a peahen.

A final theory for the tail is that it may have been used as a climbing aid. This ties back to the possibility of Leaellynasaura being arboreal. One can easily imagine Leaellynasaura with a prehensile tail. The arms of Leaellynasaura are not known, but they could have been longer than in other elasmarians, if the animal was living an arboreal lifestyle, maybe tipped with opposable thumbs like Hypsilophodon. And even if they weren’t, it doesn’t mean Leaellynasaura couldn’t have been arboreal; anteaters and tree kangaroos don’t have very long primate-esque arms, but they’re arboreal anyway.
I want to be clear- this is highly speculative. There is currently no direct evidence suggesting Leaellynasaura was arboreal, and there’s just as much evidence against the idea- Leaellynasaura had very long legs, which is a classic cursorial adaptation, and the tail’s flexibility could just as easily have been used as a rudder, allowing the animal to pull insanely sharp turns when running away from predators. We’re speculating based on what little we know about the animal’s anatomy. It’s important to think about dinosaur appearance and behavior in terms of informed speculation, because we don’t know how they actually lived and for the most part we never will. The tail could have been used for any one of these purposes, or all of them, or some combination of them, or maybe none of them. We can let our imaginations run wild wondering what it was used for.

Dig A Tunnel
Regardless of whether or not Leaellynasaura spent some of its time in the trees, it appears to have slept underground. Along the rugged Otway Coast in southern Victoria, three fossilized dinosaur burrows were discovered in 2009. These burrows each stretched about 2.1 meters underground, gently descending to a wide terminal chamber. The tunnels all curve sharply at almost a right-angle, and then immediately curve again, rather like a grease trap. They’re known to be burrows because they were infilled with sediments from a flood later on that doesn’t match the sediment they were originally dug out of.
These burrows were just the right size for small ornithopods to have resided in. They were too big for the mammals of the time, and much too small for the local theropods. No skeletal remains were found within the burrows, so they can’t be definitively assigned to Leaellynasaura, but it can be inferred that they did belong to Leaellynasaura or a related ornithopod, because they are virtually identical to the burrows of Oryctodromeus, a related species that lived in Montana during the late Cretaceous. This Montana burrow actually had three fossilized Oryctodromeus in it, an adult and two babies. Oryctodromeus was a parksosaur, a clade closely allied to Leaellynasaura’s clade elasmaria, and in light of these discoveries it’s been hypothesized that many small ornithopods may have been burrowers.
In Antarctic Gondwana specifically, burrowing would have been a wonderful way to help small dinosaurs like Leaellynasaura survive the brutal winters. They couldn’t migrate because they were too small, so they had to hunker down where they were. Burrowing would have been an excellent way of keeping warm. Maintaining a warm temperature isn’t a small thing- platypuses in Tasmania can make their winter burrows a toasty 64°F above the outside air temperature. Where Leaellynasaura was living, well within the Antarctic Circle, maintaining a warm domicile would have meant the difference between life and death. The way the burrows curve like a grease trap would have kept heat in even more effectively, and also would have served the dual purpose of deterring the entry of predators.

Burrowing present many exciting possibilities for the behavior of Leaellynasaura. Firstly, the three burrows were all directly beside each other, which may suggest gregarious behavior, “flocks” creating tight knit burrow-colonies over winter. This isn’t unheard of. Modern puffins, which congregate in massive flocks, dig one-meter burrows for nesting. And we know certain small ornithopods, like Hypsilophodon, were herd animals because of fossil beds containing many individuals of the same species that all died together.
It’s likely these burrows were only occupied seasonally. The burrows were located in alluvial deposits, near what appears to have been a braided river valley that would have undergone seasonal flooding in spring when meltwater swelled its banks. Burrowing also would have been more difficult in the dark of winter, when the ground was frozen solid, and burrow depth would have been limited by permafrost- modern tortoises and puffins will stop digging when they encounter permafrost, or even choose to dig elsewhere. So it can be inferred that the burrows were probably dug in autumn, before the first frost, and abandoned in spring at first thaw. Burrows would have been filled, collapsed, and eroded away on a seasonal basis, which is why these three are so exceptional- they were dug well enough to be infilled, rather than simply collapsing like most others.
The burrows may also have been used for nesting- they certainly appear to have been by Oryctodromeus- thus protecting eggs and/or hatchlings from the elements and predators. If this is the case, a potential breeding rhythm for Leaellynasaura presents itself- a summer rut, with new couples bedding down for winter to incubate their eggs in the warm protection of the burrow, and introducing the hatchlings to the world in spring. This of course is highly speculative, and unless we find Leaellynasaura eggs in a burrow in the future- highly unlikely, given the dearth of dinosaur fossils in Australia- we may never know.
It should be noted here that, due to the fragmentary nature of Leaellynasaura fossils, we don’t know if it had any fossorial (burrowing) adaptations. Oryctodromeus did- it possessed a fused premaxilla (a snout modified for shoveling), attachment points on its scapula for strong digging muscles in the arms, and a modified hip that would have been helpful for bracing the body when burrowing with the forelimbs. These are all things that it would make sense for Leaellynasaura to have, but we simply don’t know if it did or not. Regardless, it’s reasonable to conclude that it and the other small ornithopods it coexisted with probably burrowed to survive in winter.

And yet, Leaellynasaura doesn’t appear to have hibernated. Like most animals, dinosaurs laid down annual rings in their bones, not dissimilar to the growth rings of trees. This enables paleontologists to have some understanding of dinosaur growth rate and how long they may have lived. The leg bones of Leaellynasaura and other Australian elasmarians have been analyzed and they show no signs of Lines of Arrested Growth (LAGs). LAGs are a specific type of ring that appears on bones in hard metabolic times, when the animals aren’t eating a lot and thus aren’t able to lay down as much bone tissue. LAGs would be expected to appear if an animal is hibernating or is suffering from a shortage of food. For example, brown bears show LAGs when they hibernate, and polar bears, which no not hibernate, nevertheless show LAGs in winter, when food is scarce. The lack of LAGs in Leaellynasaura (say that three times fast) suggest that not only was it active in winter, it fared rather well despite the cold. This also strongly suggests that Leaellynasaura was warm-blooded, able to regulate its own internal body temperature. Dinosaurs in general are now thought to have been warm-blooded, far from the stupid, ectothermic swamp-dwellers of the mid-1900s, and polar dinosaurs especially would have needed to be endothermic to survive the long dark of winter.
Interestingly, the study also looked at fragments of a leg from a Victorian theropod called Timimus. Timimus is known from even more fragmentary material than Leaellynasaura- all we have is the leg. Two legs, to be precise, one from an adult and one from a juvenile, found right beside each other. Paleontologists aren’t sure what it was- it may have been an ornithomimid (“Galli-uh-Galla-Gallimimus”), a dromaeosaur, or even a tyrannosauroid. Whatever it was, it seems to have hibernated during the winter because its bones do exhibit LAGs.
Finally, this analysis of bone histology enabled researchers to figure out the life cycle of Leaellynasaura and other small Victorian ornithopods. They grew very quickly in the first two to three years of their lives, reaching skeletal (and possibly sexual) maturity between the ages of five and seven, and then continued to grow at a slower but steady rate for the rest of their lives.
Assignment Issues
Now, I have to say this- because Leaellynasaura is known from such fragmentary material, there has been debate over whether or not all of the bones assigned to it are actually from the same species. In 2013, Matthew Herne analyzed all known fossils assigned to Leaellynasaura, and found that he couldn’t confidently assign any postcranial material to the species. Only the original skull holotype and a few other skull elements are for sure known to have belonged to Leaellynasaura. The legs and super long tail are currently in limbo. But, despite this, we know that a small ornithopod from early Cretaceous Australia had a super long tail. The tail fossil is from a single individual, which preserved the entire tail, but it didn’t include anything past the thoracic vertebrae.

Personally, I don’t view this as a particularly big issue- yes, the assignment is uncertain, but only because it’s impossible to assign the tail of an animal to its skull without knowing anything about what was in between. The fact remains- there was a small ornithopod with large eyes and a large brain, and a small ornithopod with a phenomenally long, flexible tail. There are other small ornithopods known to have lived concurrently to Leaellynasaura- Atlascopcosaurus, Qantassaurus, and Diluvicursor. None of them are likely candidates for ownership of the tail. Atlascopcosaurus was too large to be the owner, Qantassaurus appears to have been too stocky, and Diluvicursor’s tail is known and was much shorter and more robust than the tail assigned to Leaellynasaura. Barring the existence of a fifth species of ornithopod- which is entirely possible- it still seems reasonable to portray Leaellynasaura using all the material that has been assigned to it. It’s much cuter this way, at any rate.
I’ve only focused on Leaellynasaura in this piece, but rest assured, it did not live a lonely existence. In part two, we’ll be discussing some of the dinosaurs- and other odd creatures as well- that Leaellynasaura shared its world with. Stay tuned!

Source
Martin, A. J. (2009). Dinosaur burrows in the Otway Group (Albian) of Victoria, Australia, and their relation to Cretaceous polar environments
Wikipedia page on Leaellynasaura
Continent of Curiosities: A Journey Through Australian Natural History by Danielle Clode, 2006
Sharpe, et al (2017). Revealing the skeleton of the polar dinosaur Leaellynasaura amicagraphica using synchrotron computed tomography
Chinsamy, et al (1998). Polar Dinosaur Bone Histology
Woodward, et al (2011). Growth Dynamics of Australia's Polar Dinosaurs
Woodward, et al (2018). The bone microstructure of polar “hypsilophodontid” dinosaurs from Victoria, Australia
Bell, et al (2018). Ornithopod diversity in the Griman Creek Formation (Cenomanian), New South Wales, Australia
Kitchener, et al (2019). High-latitude neonate and perinate ornithopods from the mid-Cretaceous of southeastern Australia
Herne, Matthew (2009). Postcranial osteology of Leaellynasaura amicagraphica (Dinosauria, Ornithischia) from the early Cretaceous of southeastern Australia